
EDITORIAL PREFACE
This chapter and the two that follow employ the trans-temporal approach to adult sexuality, as defined in Chapter 1, by tracing the way in which males and females differentiate from the beginning of embryonic development to birth, and then during infancy and childhood.
Because this same method is used, a certain degree of redundancy is inevitable. Chapters 2 and 3 run about the same course as long as they are dealing with the facts of prenatal sexual differentiation, and some question may arise as to why both authors have dealt with the same evidence. However, closer analysis reveals differences in interpretation which presage even wider theoretical differences to come.
Milton Diamond’s interpretive bias is heavily physiological, placing very strong emphasis on prenatally organized sex differences in the brain as determinants of behavioral differences throughout life. John Money’s formulation also strongly implicates biological control of sex differentiation during very early development, but he lays primary responsibility for the final determination of psychosexual differences upon social conditioning from infancy onward. It has been judged desirable to provide a forum for both points of view, since many of the unsolved issues demand as catholic an approach as can be presented.
According to Diamond’s hypothesis, sex differences in the brain are established very early, under the influence of genetic and hormonal factors, so that within 4 to 5 weeks after conception “crude neural programs are organized which eventually will mediate the individual’s reproductive and sexual patterns.” Diamond is quite explicit in defining just what these “patterns” may be, and in fact structures his entire theory in terms of them. A person’s sexual patterns are his or her ways of behaving which reveal maleness (aggressiveness, assertiveness, et cetera) or femaleness (subtlety, passivity, dependency, et cetera). Sexual identity refers to each individual’s self-perception as a male or a female. Sexual object choice is the sex of persons preferred as “erotically interacting partners,” and sexual mechanisms are the physiological responses involved in performance of the male or female role during physical sex acts.
Each of the foregoing patterns depends upon separate brain circuits or “neural programs.” Male and female programs are laid down in both sexes; but masculine mechanisms are more strongly developed in genetic males and feminine mechanisms supervene in genetic females. Because of these inborn differences in reactive capacity, very young boys and girls are likely to show different responses to the same treatment. Thisprediction is in accord with some of the observations made by Jerome Kagan in Chapter 4. The concept of a basic but unbalanced bisexuality in the brain is set forth also in Money’s discussion of human hermaphroditism and in my own chapter on animal sexual behavior.
In Chapter 5, William Davenport stresses those sex differences in behavior which seem to occur in most, if not all, human societies; and Diamond asserts that in all cultures there are sex differences throughout childhood that “reflect a basic sexual organization which is constitutionally based.” At this level of generality, there might be appreciable agreement among authorities, but Diamond’s views grow more controversial as he becomes more specific, in statements such as the following:
The effects [of hormonally induced brain organization] on sexual behavior … [are] believed to [include] erotic response levels, arousability, genital mechanisms, sexual identity, and sex-related biases in the spontaneous initiation or acceptance of various activities, as well as choice of sexual objects.
A few other points concerning Diamond’s theoretical point of view should be mentioned. Although there are frequent references to the importance of environmental determinants of sexuality, their principal role appears to be seen as one of evoking the expression of behavioral characteristics which were “biologically programmed” earlier in development.
In addition to the chapters by Money and by Kagan, there are others which deal with determinants of sex differences and which can profitably be compared with Diamond’s treatment. In particular, Chapter 9 points up the difference between masculinization and defeminization in females, and between feminization and demasculinization in males. Chapter 11 further develops the concept of bisexuality in brain organization and relates it to the occurrence of homosexual behavior in nonhuman species.
INTRODUCTION
We must bear in mind that some day all our provisional formulations in psychology will have to be based on an organic foundation. It will then probably be seen that it is special chemical substances and processes which achieve the effects of sexuality and the perpetuation of individual life in the life of the species.
Sigmund Freud, as translated by Sandor Rado
The obvious and manifest differences in physical appearance and behavior of the typical adult human male and female prompt inquiry concerning the factors which produce these differences. Some argue that physical differences are brought about primarily by biological factors, whereas learning and cultural conditions and forces are responsible for behavioral differences. On the other hand, there are others who attribute all differences to biological causes. The facts seem to indicate that both biology and environment are intimately involved in producing both the morphological and behavior characteristics. The present chapter will deal primarily with the biological factors which contribute to the differences or similarities in sexuality between human males and females. Of primary concern will be those periods of development during which primary sexual differentiation occurs and then becomes manifest.
At the outset, it should be made clear how biology, environment and culture must all be considered in the overall picture of human sexual differentiation and development. Firstly, biology acts to set potentials, limits and response biases in each individual. These establish the parameters within which culture and environment can exert their influence. From the moment of fertilization, the individual has a sex genotype bias which is constitutional and which operates to modify the responsiveness of all tissue to subsequent influences. It is the basic premise of this chapter that early biological influences affect subsequent behavioral development. For example, while steroids can affect all developing individuals, much lower doses of a male hormone are needed to induce growth of male structures and development of male behavior patterns in males than in females. Similarly, development of masculine body structures and behaviors is more sensitive to the lack of androgen in males than is the case in females. Thus, in considering behavior, two individuals of different sex with equally high motivation to be good swimmers, musicians, or lovers might differ greatly in their inherent abilities, even though they both practice equally hard and long.1 The constitutional makeup which they bring to the swimming pool, concert hall, or bedroom is different. Furthermore, interaction between the individual and the environment is two-way. Not only does the environment work on the individual to mold and shape his or her physical and behavioral character, but the individual, biased organically, works on the social and physical environment in a mutual exchange of effect. Two individuals of different size or behavioral attitude will be treated differently in the same situation by the same people. All that is said in the rest of this chapter should be appreciated within the context of these reciprocal interactions.
In any discussion of sexuality, it is extremely important to realize that definitions of male or female (even for the description of traits) are often affected by retrospective judgments involved in establishing the original categories. For example, the conclusion that roundness or softness are feminine traits, whereas angularity and hardness are masculine, is a judgment based on the findings that most males have physical features which can be categorized as hard and angular and related to muscular activity, and, in contrast, most females have characteristics classifiable as soft and round and which can be related to childbearing and nurturance.
While these generalizations hold true for most populations, the criteria may not apply to any specific individual. The generality is valid, nevertheless. Another example would be the decision as to whether mounting behavior or aggressive sexuality should be considered masculine, while being mounted or being sexually submissive should be regarded as feminine. We certainly can choose items that will reflect such sex differences, but the items of choice, while they may be reality-oriented, are idiosyncratic and may be subject to contrary opinion. Finally, we can consider subtle observer bias in the definition of male versus female behavioral characteristics. Masculinity or femininity of an individual may be considered reflected in the choice of an out-of-the-home career or preference for a domestic role. Choice of a career might be considered an indication of masculinity. In addition, the desire to have, or spend time with, children might be considered an index of femininity. These, however, are observer-determined retrospective categories of choice, based upon what our culture usually regards as the end product of typical male or female development. One must always be mindful of the fact that it is the culmination of all interacting forces to which we attend. Therefore, attributing a particular facet of behavior or morphology to biological innateness or environmental determinism, while it may be true in fact, can be difficult to document. As a generality, without other data, it is probably best to consider traits as inherently male or female in humans if they follow logically from what we can expect, on an evolutionary continuum, as revealed in the behavior of males and females of many other mammalian, and primate species. To this, we of .course add the flexibility and adaptability which are highly developed human characteristics. When humans are considered to deviate from what we might consider normal in evolution, we should be skeptical.
DIFFERENTIATION


The complexity of the variables involved in human development allow for an infinite variety of inputs into any individual. Starting from the genetic inheritance involved when sperm and egg meet at zygote formation, a progression is seen between simultaneous and sequential influences, Initially these are mainly genetic and endocrine forces which affect the development of the gonads, the external and internal morphology, andthe nervous system. These will be seen to be related to sex assignment, sex rearing, psychological bias, gender roles, behavioral patterns, sexual identity, sex object choice, sexual mechanism, and other concerns during childhood, puberty and adulthood.
Genetic and endocrine factors, while always affecting the individual, may be considered to exert their primary influence during three biological periods: predifferentiation, differentiation and development. The first period biases the outcome of the second and the second influences the third. These periods differ in duration and significance. The particular influential features within each period can be categorized as follows:
- Predifferentiation
- Genetics
Gonads
Endocrines - Differentiation
- Endocrines
Internal morphology
External morphology
Nervous system - Development
- Internal morphology
External morphology
Nervous system - Sex assignment and rearing
Psychological sex (sexual patterns and mechanisms, sexual self identity, sex object choice)
Puberty
Adulthood
The predifferentiation stage is the initial period of ontogeny which precedes the appearances of visible differences between the sexes. During this early embryonic period (the first 7 weeks in human beings), the individual might be considered to be in a sexually indifferent stage (Fig. 2.1). The lack of visible distinction between the sexes leads to the common application of such terms as “neuter” or “indifferent” to these stages, or to the organs and systems under discussion. It should be understood that this refers only to appearances (phenotype)—the potential competence of the tissues differs markedly between the sexes and also among individuals. Competence is the ability of a tissue to react to a particular developmental stimulus. At this point in ontogeny, the genital primordia maintain a bisexual potential to develop along either male or female lines. This bisexual potential does not necessarily involve an equal capability to be male or female in every individual, nor are the tissues equally competent to be masculine or feminine. From the time of zygote formation, genetic differences among individuals impose upon each person certain limitations with respect to masculine or feminine development. These limits can be approached under the influence of suitable stimuli, but in normal humans cannot be transcended by subsequent biological or environmental factors.
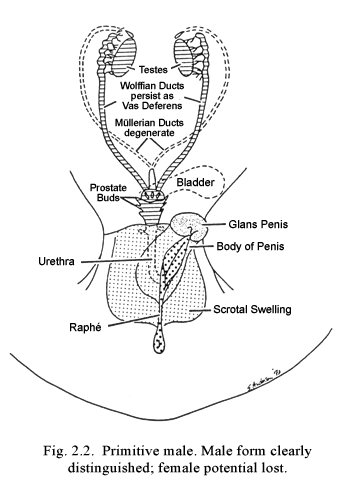
The second period of interest is that of sexual differentiation, which occurs during the 4- to 5-week period following predifferentiation. During this interval, the individual loses a great deal of his or her relative pansexual potentiality and progresses definitively toward a predominantly male or female existence. At the end of this stage, the individual might accurately be described as a “primitive” male or “primitive” female (Figs. 2.2 and 2.3). The internal reproductive organs are concordant with the genetic sexof the individual and the external sexual organs are appropriate to his or her genetic and endocrine nature. In addition, crude neural programs that eventually will mediate the individual’s reproductive and sexual patterns are organized.
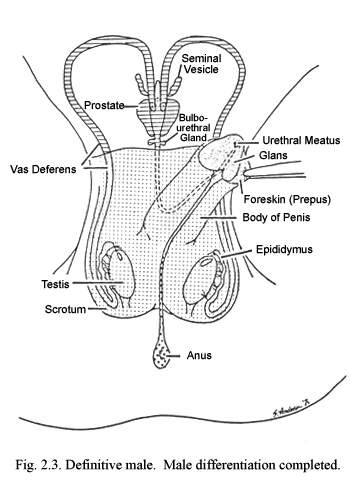
Subsequent to the period of sexual differentiation occurs the much longer and diversely influenced phase which goes by the all-inclusive term of development. Due to its complexity, the protracted time span involved in development can be subdivided into post-differentiation gestational, prepubertal, pubertal and postpubertal, and mature intervals. It is during the post-gestational periods that the individual matures biologically (Figs. 2.4 and 2.5) and psychologically. It is also during this period that the person is most influenced by environmental forces, although strong biological factors remain nevertheless crucial. This chapter will concentrate on those periods preceding puberty.
Predifferentiation
From the time of fertilization, the genetic stamp of the individual is established. As cell division progresses, this mark is transferred to every cell of the body. Typically, the mark reflects either a female XX constitution or a male XY constitution. In human embryos, the effect of this heritage usually does not become manifest until the sixth or seventh week in utero. Prior to this time, the human fetus can be regarded as appearing neuter or indifferent. It is indifferent in internal and external appearance (phenotype.), containing neither testes nor ovaries, but only undeveloped gonads located near the kidneys.
At this stage of development, two systems of primitive ducts exist in the fetus: One is the Müllerian duct system with the potential to give rise to oviducts, uterus, and upper vagina; the other is the Wolffian duct system, which can give rise to the male epididymides, vas deferens, and seminal vesicles. Despite the simultaneous existence of both duct systems and the apparently indifferent gonads, the genotype has pre-established inherent male and female potentialities in these embryonic structures. The inductive nature of the genotype is such that the indifferent gonads will develop into ovaries in individuals containing two X chromosomes, whereas the gonads will normally develop into testes in individuals bearing a Y chromosome. In the human male and female, chromosomes are not balanced biologically, as they are in several other species, nor is there, at this stage, an overriding male differentiating influence from the autosomes. If a single X chromosome is present, gonadal agenesis or dysgenesis will result. If an individual’s genetic constitution includes any combination of more than two sex chromosomes, the presence of a single Y chromosome will insure the development of testes, regardless of the number of X chromosomes present. The presence of more than two sex chromosomes does not impart any sexual advantage to an individual; in fact, the opposite is more likely to occur. Most such individuals are sterile and, regardless of their upbringing, display low libidinal levels. It is instructive to note that individuals with genetic constitutions other than XX or XY are not uncommon in human beings, occurring in as many as one in 100 or 200 persons.
The manner in which chromosomal factors influence the direction of development of the gonad is not yet clear, but it seems apparent that the male and female potentialities in the gonad are pre-established. in the medullary and cortical gonadal components at a very early stage. This seems obvious, since differentiation of the gonad into an ovary or testis involves the gradual predominance of either the medullary or cortical component over the other, rather than the transformation of one sex element into the other.
In an individual with a Y chromosome, the medulla develops so that it envelops developing germ cells, which will become the future spermatogonia. In the individual containing two X chromosomes, the medulla forms in a circular pattern, excluding the germ cells from their midst and pushing them to the periphery. These germ cells will give rise to the ova. Thus the relative distribution of cortex and medulla develops, with the allocation of germ cells establishing the prototype for the future ovary or testis. The sex genotype is considered to act primarily in initiating sexual differentiation by determining the developmental pattern to structure the gonad. It is appropriate here to define differentiation. This refers to the developmental process responsible for intrinsic and irreversible differences in form and function among tissues. Differentiation, in this sense, is common during embryonic and fetal growth, and is uncommon in the adult. Differentiation must be contrasted with modulation, which is more common in the adult. Modulation refers to temporary and often reversible changes which occur in response to different stimuli and different environmental conditions.
Considering this histological differentiation of the gonad, we may say that, after genetic sex determination at fertilization, the differentiation of the gonad isthe most basic event in sexual development. The type of gonad—ovary or testis—possessed by an individual is defined as his or her primary sex characteristic.
Sexual Differentiation
Sexual differentiation occurs in analogous ways in the genitalia and in the nervous system.
Genitalia. Once a definite male or female gonad differentiates, it, in turn, gives direction to the development of the two coexisting pairs of genital duct structures. If the gonad differentiates as a testis, it produces an as yet unidentified hormonal substance which has the capability of simultaneously inducing masculine development of the Wolffian duct system and inhibiting development of the Müllerian duct system (Fig. 2.1). This formative influence exerted by one tissue (or substance) on a neighboring tissue is called induction, and it results simultaneously in two types of determination. Induction insures the continued or developed presence of a particular tissue (evocation determination), and at the same time it determines the characteristics possessed by that tissue (individuation determination). The significance of specifying the dual nature of this determination will be enlarged upon below.
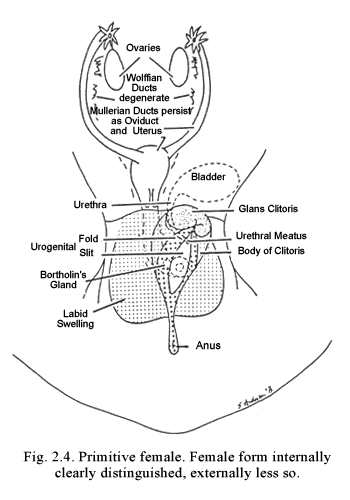
Since the biochemical nature of the inductor substance is unknown, I have for convenience termed it “fetal morphogenic testicular substance” (FMTS). The name serves to define the period of activity and the source and primary level of action of the hormone or hormones involved. If the gonad differentiates as an ovary, no substance corresponding to FMTS (i.e., acting to induce development of the Müllerian duct system and regression of the Wolffian duct system) appears.2 In crucial distinction from the case in the male, where the presence of FMTS is required for normal differentiation, in the female, it is the absence of FMTS that results in preservation of the Müllerian duct system and regression of the Wolffian ducts (Fig. 2.3). This finding is corroborated by results of animal experiments and by clinical experiences, which show that fetal castration, gonadal destruction, or absence of gonads prior to differentiation invariably result in sexual development along female lines.. The Wolffian ducts regress and the Müllerian duct derivatives become dominant. If differentiation occurs along male lines, the major portion of the Wolffian duct system gives rise to the vas deferens and also portions of the epididymis, common ejaculatory duct, seminal vesicle and trigone of the bladder. If differentiation proceeds along female lines, the Müllerian ducts persist as the oviducts, uterus and upper vagina.
Simultaneously with differentiation of these genital reproductive structures, sexual differentiation of other common urogenital structures takes place. For example, the common urethral glands, and glands of the urogenital sinus seen in the indifferent stage, give rise to the prostate gland in the male and urethral and Skene’s glands in the female.
The significance of the evocation-determining properties of an inducing substance like FMTS is thus apparent; a structure is physically maintained and its existence, or that of its derivatives, is assured. Those tissues that are not evoked, regress. The significance of individuation determination is that a tissue is given a certain sexual bias, so that it maintains permanently a modified responsiveness to future stimuli. For example, Wolffian duct derivatives such as the vas deferens are more responsive to androgens than to other steroids, whereas Müllerian duct derivatives such as the uterus are always more responsive to estrogens.
Following differentiation of the internal genitalia, analogous processes come into play, promoting sexual differentiation of the external genitalia. Prior to sexual differentiation, a seemingly indifferent genital tubercle and pair of genital swellings exist (Fig. 2.1). In human embryos, these genital structures remain phenotypically undifferentiated until approximately the end of the third month of fetal life. This long interval prior to differentiation of the external genitalia, in comparison with the earlier development of the internal genitalia, is believed to he an important factor accounting for the frequency and wide variety of genital incongruities seen in human. Whereas differentiation of the internal genitalia along male or female lines is dependent upon FMTS, this substance is not crucial for differentiation of the external genitalia. Instead, sexual development of the external genital structures is mainly dependent upon the presence or absence of testicular androgens, such as testosterone. The tubercle, genital swellings and associated structures differentiate as female genitalia in the absence of male hormones. While usually in the testes, the origin of the androgens to which the external genitalia can respond is immaterial: testicular, adrenal. maternal or exogenous androgens all are effective. This is a crucial point, since there are many clinical conditions in which the presence of nontesticular androgenic substances induce masculinization of a female. For example, an overactive fetal or maternal adrenal may produce androgens at an appropriate time in development, and under such conditions that the female fetus will develop virilized genitalia (adrenogenital syndrome). Aberrantly functioning tumors of the maternal ovary also can masculinize the female fetus (arhenoblastoma). Still another cause of such masculinization is seen when pregnant women take, or are given, androgenic substances (drug-induced masculinization).
Normally the newly developed testes produce the androgens necessary for male development. In the male (or female) with androgens, the genital tubercle elongates and incorporates the urethral groove so that an imbedded, tubular penile urethra develops within a cylindrical phallus—the penis. The genital swellings enlarge to form the scrotal pouch, which eventually will contain the testes, when they descend from their position near the kidneys (Fig. 2.4). During the predifferentiation phase, conditions and appearances of the external genitalia of the female are essentially like those of the male except that the urethral groove is shorter. In the absence of androgens, the urethral folds flanking the urethral groove do not fuse, but persist as the labia minora, flanking the urogenital vestibule. Instead of pouching, the genital swellings grow together between the anus and the urogenital vestibule. Their point of fusion becomes the posterior commissure and the lateral portions grow as labial swellings to persist as labia majora. The lower vagina develops concomitantly with formation of the urogenital vestibule. Estrogens are not crucial for differentiation of the external genitalia of either sex (Fig. 2.5).
While necessary for differentiation of masculine genitalia, the presence of circulating androgens is not by itself sufficient for masculinization. The capability for the appropriate end organ response of the pertinent tissues is required. By virtue of his genetic inheritance, the male with a normal XY genetic component typically has tissues which are more responsive to androgen than does the female with her normal XX genetic makeup. However, various specific genetic conditions can preclude or alter this natural responsiveness to androgens. An extreme example is seen in some males who, despite an XY chromosomal complement, and despite the presence of testes and normal levels of androgen production, have the appearance of females. An inherited inability of their tissues to respond at all to these androgens causes these males to develop phenotypically as females, and they are called “testicular females” or “androgen-insensitive males.” Since differentiation of the internal genitalia depends upon FMTS, and not upon androgen, these individuals have internal genitalia which are typically male. This would be a case of the internal genitalia having been evocated as a normal male without normal male individuation determination of external or other tissues. This example illustrates a crucial point. An individual’s genetic heritage remains crucial throughout life for psychosexual and morphosexual development.
In the 1940s and 1950s, human sexual differentiation was conceptualized as depending upon both male and female evocator substances. Male differentiation was thought to be dependent upon androgens and female differentiation upon estrogens. For the human, this theoretical framework has been supplanted by the present concept of only FMTS and androgens being necessary for development along male lines. The absence of FMTS and androgens is necessary for development along female lines. The magnitude of effect of FMTS and androgens is dependent upon genetic heritage.
Period of maximum sensitivity (critical periods). Heretofore, we considered various inducer substances and their effects on various tissues. It must be emphasized that the ability of tissues to respond varies with time. This responsiveness is dependent upon the time of onset, duration and magnitude of exposure to the inducing substance. The critical nature of these time relations is known best from experimental studies, but some clinical data are also available. In general, for humans, it seems that the timing involved with the many facets of sexual differentiation and development are only slightly known. For morphological characteristics, however, most organization occurs between the 7th and 12th week of the gestation.3
Sexual differentiation of the neural tissues. In addition to genital tissues, sexual differentiation of a different type is also directly related to our interest. This involves primarily sexual differentiation of the neural tissues and the neural-endocrine axis associated with reproduction and sexual behavior. Present evidence indicates that processes somewhat analogous to the hormonal mechanisms involved in genital differentiation are also active in differentiation of the behavior-mediating and reproduction-mediating neural tissues. But, in terms of time, these neural tissues are the last to differentiate, the process probably continuing until after birth. Sexual differentiation of the internal and external genitalia is apparent prenatally; sexual differentiation of neural tissues in general is initiated prenatally, but is not apparent until after birth and is especially vivid after puberty. The concept of neural tissues being programmed prenatally for later manifestations recalls the genital tissue response of individuation to evoking substances. Emphasis is focused on neural development, since all behavior is invariably a neural function.
In the adult individual, this neural organization will affect reproduction mainly via gonadal, and, in the female, genital tract cyclicity. In the human male, cyclic activity is not seen and spermatogenesis is the reproductive process affected.4 The effect on sexual behavior is on a broad plain and believed to influence: erotic response levels, arousability, genital mechanisms, sexual identity, and sex-related biases in the spontaneous initiation or acceptance of various activities, as well as choice of sexual objects. The analogy between differentiation of the genitalia and neural tissues must be employed with caution and therefore will be discussed in detail.
The development of the neural tissues is much more complicated than that of the internal and external genitalia. The genitalia are usually considered functional or appropriate in simple, male-female categories, without conflict. Differentiation of the genitalia is relatively obvious and once it has occurred, future development involves little more than growth and functional activation. For the neural tissues, whose capability and functioning are revealed only after birth, and in some cases only after puberty, a comparable, stable male-female dichotomy is observable, mainly for purely reproductive functions such as gamete production, the menstrual cycle, pregnancy and lactation. An individual may be stimulated by hypothalamic-pituitary influences to show either cyclic ovulation and menstruation, or tonic spermatogenesis, but not both simultaneously. For differentiation of the genital tissues along male or female lines, fairly unidimensional processes are involved. A genital tubercle will develop as a penis or a clitoris, but never both simultaneously. Similarly, the Müllerian duct system will either survive or regress reciprocally with the Wolffian duct system. In the mature individual, both do not usually coexist. Thus, the functioning of the internal genital tissues is like that of the neural tissues for the reproductive obligations; the individual is either male or female.
In the case of behavior-mediating neural tissues, the male or female dichotomy is not clearly distinguishable. Stereotypes of male and female behavior exist, but the observable points to be considered are multiple and their definitions often superficial. Also, sexual behavior is modifiable postnatally while gonadal and genital functions typically are not. Indeed, facets of sexual behavior are only finally differentiated during the processes of maturation. When behavior-mediating neural tissues are considered, behavior patterns considered male and female may exist simultaneously in the same individual and may be manifest at various times to a greater or lesser degree. This notwithstanding, development of the genital tracts and external genitalia does have its counterpart in endocrine effects upon development of the behavior-mediating neural tissues. In males, the presence of androgens is crucial for development of normal male sexual behavior patterns and mechanisms. The absence of androgens seems to impede or completely prevent such development and simultaneously to facilitate the emergence of feminine patterns and mechanisms—this, despite the style of male upbringing. In females, androgens present during the period of differentiation to some degree foster the development of masculine behavior and simultaneously inhibit behaviors more typically female—this, also, despite female upbringing. The magnitude of the effect produced by the presence or absence of sexually differentiating hormones is dependent upon the time of onset, duration and degree of endocrine exposure on the one hand, and the responsiveness of the androgen labile tissues on the other hand. There can be no doubt, however, that males are usually more responsive to androgens than females at any time of development, and similarly females are more responsive than males to the absence of androgens in their development. With this caution, we thus should not anticipate androgenized females to be comparable to males or androgen-deficient males to be comparable to normal females.
This distinction between the factors determining sexual differentiation of the external and internal genital tissues and those affecting differentiation and organization of the behavioral mediating neural tissues is significant. In the case of the external genitalia, it is legitimate to regard male and female as representing ends of a continuum, where the development of the genital tubercle toward the phenotype of one sex precludes simultaneous development toward the other sex. This can be called mutually exclusive differentiation. An example is seen in the case of phallus length. An individual cannot simultaneously have a large penis and a small clitoris. Similarly, when one of the two sets of internal genital duct systems differentiates, one becomes dominant and the other regresses; usually to vestigial status. This can be termed simultaneously exclusive differentiation. For the neural tissues mediating reproduction and sexual behavior, we have at present no way of knowing whether one or two, or more sets of tissues must differentiate. Experimental and clinical data, however, lead to the belief that the neural tissues differentiate in yet a third manner, with characteristics of both internal and external genitalia. It is probable that differentiation of the neural tissues involved in procreational processes, like the external genitalia, is mutually exclusive in character. The individual can show hypothalamic-pituitary-gonadal processes which are either tonic or cyclic, but not both. In the presence of androgens, the neural tissues involved in procreation are organized along male lines, i.e., a tonic, non-cyclic, hypothalamic-pituitary relation will obtain. In the absence of androgens, a female, cyclic type of control will develop.
Human reproductive capacity is characterized by its cyclic nature in females and its tonic nature in males. For example, the cyclic process of ovulation, uterus preparation and menstruation, requires a complexity of neuro-endocrine coordination which is provided only by a female hypothalamic (nervous system) differentiation. Transplants of ovaries and uteri to a male do not show cyclic activity despite adequate vascular and histo-immunological acceptance by the host tissues. Spermatogenesis in the male, while a cyclic process in itself, is dependent upon relatively tonic hypothalamic-gonadotropic stimuli.
The neural tissues involved in sexual behavior, like those of the external genitalia, respond to testoids. In contrast with those involved in procreation, however, they differentiate somewhat like the internal genitalia, but in a manner of differentiation that might be considered simultaneous but not exclusive. Behavior patterns, biases and mechanisms of one sex will be dominant, but those of the other sex may regress only slightly and remain subordinant or latent (subdominant). The determination of which are dominant behaviors and which subordinant depends upon threshold levels of response or response probabilities (biases) for the behaviors, and must be considered in terms of evoking or suppressing stimuli. Behavioral tissues thus form a unique category, since male and female patterns and biases may coexist and, depending upon how they are evaluated, appear equally well developed simultaneously. The mode of differentiation, initiated now, will only become apparent much after birth. For example, one individual may show a good deal of sexual aggressiveness and promiscuity in initiating sexual contact (considered a male trait), and at the same time exhibit a great deal of passivity or submissiveness when engaging in coitus (considered a female trait). Without clearly defining the actual behavioral traits affected by genetic and endocrine factors, it can be said that they probably are invested with limits and biases influencing the direction and emphasis that will be mediated by crucial neural tissues and later may be affected by culture. These limits need not lie upon a simple male-female continuum. The behavioral repertoires of males and females can be considered parallel and discontinuous rather than continuous, and thus they should not be subject to the same evaluation criteria as applied to morphology. An additional, complicating, factor is that behavioral sexual differentiation and development most profitably must be evaluated on at least four levels. Usually these are in concert, but their development need not be.
Differentiation along four levels of behavioral sexuality. The four levels of sexuality under consideration will be presented here in their adult mode to enable the reader to follow their trend of development. It is assumed that the neural tissues involved are individuated pre- and neonatally in development. With varying degrees of modifiability, they will continue to develop throughout life. In addition to these levels, others may be distinguished, but these basic four are considered a minimum to provide a sexual profile for an individual.5
The first manifestation of sexual behavior can be assessed by sexual patterns, e.g., for maleness: direct aggressivity, assertiveness, large muscle motor activity; and for femaleness: subtlety, passivity and dependency, nurturance, small muscle motor activity. Patterns are largely susceptible to cultural and learning factors. The second manifestation of behavior is sexual identity, i.e., the sex to which an individual ascribes: male or female. This feeling of identity is typically reinforced by society, but can exist independently of society’s inputs. The third is sexual object choice, i.e., the sex of the individual chosen as an erotically interacting partner, usually a male for a female, and vice versa. This, too, is typically reinforced by society, but can exist independently of its support. The fourth assessment of sexual behavior involves sexual mechanisms, e.g., those features of sexual expression over which the individual has little conscious control: the ability of a male to get and maintain an erection, have an orgasm and/or ejaculate; the ability of a female’s vagina to lubricate and for her to be orgastic.
In most individuals, these four characteristics will be in concert among themselves and with other criteria of sex. This is to say that normal genetic males or females will develop the appropriate sexual anatomy, will see themselves as males or females, and will eventually act erotically and characteristically with an opposite-sexed partner. It is hypothesized that closely related and interacting, but different, sets of neural tissues are involved in pattern, identity, object choice and mechanism functioning, as there is a separate set for reproduction. While there is as yet little anatomical evidence that separate neural tissues exist for each of these components of sexual behavior, clinical considerations will be presented below to indicate that indeed quite distinct tissues might develop and exist. I believe most difficulties in interpreting or discussing either developing or existing adult sexuality and gender characteristics. similarities as well as differences, is that these different levels of behavior are often mistakenly discussed along one male-female continuum despite the disparate nature of the traits under consideration.
Each set of tissues theoretically should have its own critical period for differentiation, and should develop in its own mode. Due to various situations, a distortion of normal, unified development may occur. An immediate clinical example of two seemingly divergent levels coexisting in the same individuals is the female who identifies herself as a female, and is so identified by others, but who simultaneously has erotic preferences for other females (i.e., a lesbian). Clinically, this concept of different sets of neural tissues provides a framework, a method of conceptualization, within which the most vexing etiological problems of sexuality could readily be interpreted, e.g., how can individuals become homosexual, heterosexual, bisexual, transsexual, etcetera. An individual would thus be biased toward transsexuality when the tissues associated with morphology and those associated with identity differentiate out of phase with each other, so they come under different influences, one under male stimulation, the other not. An individual would be biased toward homosexuality by the neural tissues for gender identity and object choice developing out of phase with each other. Since the factors of tissue competence, developmental timing and hormone secretion dosage could vary, all grades of heterosexuality, homosexuality or transsexuality could be induced (evoked and individuated) to interact with future environmental influences.6
The effects of considering the differentiation of the neural tissues as extensively as presented here, and contrasting such differentiation with the development of the morphological tissues, allow us to entertain different expectations for the separate tissues involved. An apparently well-developed and masculine-appearing penis normally is anticipated to function as one. In contrast, there are no comparable landmarks for the behavioral mediating neural tissues. However, it is conceivable that since both the penis and neural tissues are dependent upon the presence or absence of androgen for normal development, growth of the penis might serve as a bioassay for the capacity or potential of the nervous system. In other words, an individual showing a good-sized and adequately functioning penis might be accepted as having been primed along masculine lines in regard to sexual behavior. It has been argued that deficiencies in copulatory performance due to lack of androgen during development actually reflect underdevelopment of copulatory organs. To this argument it should be added that underdevelopment of the phallus may be accompanied by underdevelopment of central neural tissues mediating masculine behavioral patterns. A different order of behavior would therefore be inevitable. The cause and effect relationship involved is still under investigation, and it cannot be gainsaid that an individual with a small penis may have sexual drives and performance levels considered normal, whereas other individuals with quite adequate genitalia may be unable or unwilling to do much sexually. These discrepancies may be due to varying rates of development between the tissues involved and their individual ability to change in response to evoking substances or modifying circumstances. This can obviously lead to “anomalous” disparities between genital structures and indices of sexual behavior, since the genitalia differentiate much earlier than the neural tissues but the various behaviors themselves may be dichotomized. It is likely that, starting from this stage of differentiation, different neural tissues develop separately for basic sexual patterns, sexual identity, bias toward sexual objects, and genital mechanisms.7
DEVELOPMENT
Subsequent to the period of sexual differentiation, the individual passes into a phase of growth. Phenotypically, this phase is noted mainly for quantitative changes and restructure. With some exceptions, this remains generally true for all sex-related structures and reproductive neural functions until puberty. In regard to sexual behavior, neural development, in contrast, continues to undergo significant qualitative changes. The entire span of development can best be described in terms of three main subperiods: a) differentiation until birth, b) birth until puberty, and c) puberty and maturity.
Development from Differentiation until Birth
After sexual differentiation, the internal and external genitalia continue to grow in size proportionate to their neighboring organs. The gonads, however, in late fetal life begin to migrate to positions other than those they occupied in the embryo. Migration of the gonads is termed descent and may be considered in two stages. The first stage, in which the gonad moves inferiorly from its position of origin toward its eventual adult location, is common to both the testis and ovary. The second stage, peculiar to the testes, involves movement to an extra-abdominal position in most mammals. This is not only functional in providing a thermal environment suitable for spermatogenesis, but the scrotal bulge may provide added erotic visual stimulation to the female and allow for added tactile stimulation for the male.
Passage of the testes from the abdominal cavity to the scrotum usually occurs during the seventh or eighth fetal month. At the time of birth, however, approximately 2 to 5 percent of human males still retain undescended or abdominal testes (cryptorchism). In the female, the ovaries, attached above the suspensory ligaments which persist, are prevented from descending out of the abdominal cavity as do the testes. The ovaries come in proximity with the superior fimbriated ends of the oviducts. Thereafter, they remain in this relative position which eventually will be crucial for capture and migration of the ovulated eggs.
In other regards, general body morphology seems not to be distinguished by sexually dichotomous characteristics. At the time of birth, individual males and females cannot be separated by somatotype without recourse to genital inspection; but that the sexes are constitutionally different during gestation cannot be denied. One indication of a major sex difference is differential fetal mortality. It is estimated by some researchers that at the time of fertilization 3 males are conceived for every 2 females. However, by the end of gestation, there remain only about 110 males for 100 females. The factors involved in these fetal deaths are unknown.
During this period of development, the external genitalia are no longer subject to the hormonal forces affecting sexual differentiation, but the penis and clitoris are modifiable to a marked degree. For example, virilization of females may occur due to late-developing adrenogenital problems. The neural tissues modifying sexual behavior also are still labile to steroidal influences and this lability probably is maintained for some time after birth.
Development from Birth until Puberty
At birth, although there is no positive gross visual method of distinguishing the sexes, aside from the genital differences, the sexes develop with different physiological capabilities and reactive potentials to environmental stimuli. It is obvious that from the moment of birth human males and females may be treated differently, either consciously or unconsciously. This differential treatment usually begins at the time of sex assignment which typically is made at birth and depends upon the appearance of the neonate’s external genitalia. Sex assignment biases the way in which society in general will interact with the individual and will anticipate his or her responses. However, the individual who is subjected to these sex-relating differences in rearing policies and is not a blank slate to be written upon. As explained previously, he or she comes to the world with constitutional, genetic and hormonally mediated, diasthetic behavioral biases and innate patterning related to all the different basic levels of sexuality. These induced influences orient the way the individual will act and in turn be treated; i.e., the individual can evoke or can be responsive only to specific types of stimuli from the environment.8
Sex-related mechanisms. Aside from morphological differences, there are inherent constitutional differences of appreciable magnitude. Within the first 28 days after birth, as a national average, about 25 percent more males than females die. This differences is even greater if we do not consider infants born below 1000 gms where any individual is at a disadvantage. The ratio of male to female deaths continues throughout life. The factors involved in these different mortality figures are yet unknown, but they obviously reflect inherent physiological sex differences. Considering that mortality is an extreme index, it is reasonable to assume that the morbidity differences between the sexes are similar, if not actually larger. These conditions of illness most probably evoke more attention given to males by mothers or other adults. In general, significant sex differences continue throughout life for the time of onset and severity of most diseases and, as is well known, for mortality.9
At birth, male and female populations may show significant physiological differences. Boys generally are 5 percent heavier and 1 to 2 percent taller than girls. These differences persist until ages 10 through 13, when the girls catch up with the boys and even pass them slightly. This change reflects the earlier prepubertal and pubertal growth characteristic of females.
The years between birth and puberty are often considered relatively noneventful in terms of physiological sexual development. This unfortunate misconception arises mainly for two reasons: 1) the pace of what is happening is slow; and 2) concentration is often focused only on reproductive functions and genital development.
Starting in late childhood, general body structure morphologically and functionally develops quite differently in the two sexes. Boys grow faster than girls for the first 6 months of life. The differences attained persist for approximately the first 4 years. At 6 years of age boys are assuming the general somatotype appropriate to adult men, with shoulders broader than hips and long, muscular chests. Their vital capacity (measure of the sustained energy output possible for an individual) is 7 percent greater than girls’ and their muscular capacity is 10 percent greater. This difference in muscular capacity exists even though the general muscle mass is similar in both sexes. From between 7 and 9 years of age, differences in muscular abilities and vital capacity between the sexes increase markedly and the boys rapidly increase their lead, even prior to puberty.
From the age of 1 year, there are significant sex differences in growth and deposition of skin and subcutaneous fat tissue. The skin and fat layer in girls is approximately 50 percent thicker than it is in boys. At the age of 7, girls begin showing signs of feminoid distribution of this fat deposition, mostly around the hips and buttocks. These differences continue to increase until puberty when they are markedly accelerated. From the age of 6, boys have bigger chests than girls the same age and from this age onward, girls have relatively broader hips than their cohorts.
Children destined to early puberty tend to be tall and to have an advanced bone age. Those children who reach puberty later tend to be short and to show retarded epiphyseal closure. However, epiphyseal development of girls is consistently advanced in comparison with that of boys, throughout childhood. Following the pubertal growth spurt, the child whose puberty is delayed will become a bigger adult than one in whom puberty occurs earlier. Thus girls most often end up significantly shorter than boys, and their pubertal growth usually occurs earlier than that of boys and is of relatively short duration.
Prenatal growth is extremely rapid. However, for the first two years after birth, a deceleration in skeletal growth and weight gain is noted. Subsequently, a steady decrease in growth is seen. A rapid acceleration in growth and weight in both boys and girls indicates the onset of pubertal changes.
Much of the physical development of the infant and child interrelates and correlates with psychological development. The infant discriminates self from non-self and knowledge and mastery of the environment come with increasing muscular coordination and sharpening of the sensory systems.
The more frequent and rapid gross body movement of boys correlates with their superior three-dimensional and abstract visual abilities and with their usual superiority over girls in meeting novel tasks and new situations. Boys seem better able to replace an old “set” with a new one for problem solving. The more sedentary activity of girls corresponds with their greater visual field dependency, better ability at rapid repetitive tasks and small motor movements. These constitutional differences in muscular ability and size are reflected in differences of behavior patterns.
Sex-related patterns. Allowing for individual differences with wide ranges, boys more often, without encouragement, choose those activities which require muscularity and stamina, e.g., running, climbing, wrestling. Girls more often select relatively less strenuous and conservative activities, e.g., hopping, jumping, playing house and mothering dolls. Boys generally choose to be more gregarious than girls and play in larger groups. They are more competitive but also better able to cooperate among themselves. Consider, for example, in no culture have girls’ games or activities evolved such as baseball (9 on a side), rugby, football, or soccer (10-12 on a side). Rather than role-modeling behavior, dependent upon social conditioning, these general behavior patterns are believed to be innate since they are consistently seen despite extreme differences in culture and environment. The culture will determine which games are played and by what rules. The general male patterns, however, dictate that physically aggressive, competitive games will be played.
From the phylogenetic point of view, it is instructive to note that comparable sexually dichotomous behavior patterns are seen in both free-ranging and laboratory housed nonhuman primates. Prepubertal rhesus males run and chase, wrestle, and climb more than prepubertal females; the females stay close to their mothers and play with babies more than do males. Male juveniles play in larger groups. The behavioral sex differences seen in humans represent consistent evolutionary trends.10
Practically all cultures do show these sex-associated patterns, yet different cultures do not have to show such similarities for the findings still to be significant. An anthropologically distinct culture may be considered to possess a relatively large genetic pool which is significantly different from other gene pools. Morphological traits, such as body and hair pigment, are genetically transmitted yet can vary between groups; so can behavioral traits and predispositions. Sexual behaviors, then, as other genetic characters and differences between cultures, can be accepted without being considered as inconsistent with a theory of genetic involvement in behavior. Several cultures thus need not manifest specific or similar sex differences any more than they need show similar skin color or hair type (both very obvious genetic traits). The fact is, however, that the overwhelming majority of independent cultures do show certain consistent sex-related behaviors; e.g., male physical aggression, leadership and dominance, and female nurturance and subtleness (see Ford & Beach, 1951). This is what would be anticipated from evolutionary considerations. These patterns have persisted as such among widespread cultures, most probably because they are adaptive.11
The mediating biophysical forces involved in demonstrated behavior differences between the sexes may include direct or indirect neural influences. Sex differences in neural functions and sexual mechanisms are apparent quite early. Girls 3 to 6 days of age, for example, have a skin conductance—used as a measure of autonomic activity—significantly higher than males. As neonates, girls never remain in regular deep sleep without occasional short, irregular startle movements. Neonatal boys, on the other hand, show comparatively long periods of restful sleep and periods of repeated rhythmic startle movements. The rapid body movements seen more often prenatally in boys than in girls persist into the neonatal period.
The cerebellum, responsible for muscle coordination and balance, develops differently in boys and girls. In girls, during the first five postnatal months, the cerebellum increases in size and weight about 100 percent, whereas during the same period the increase is only about 60 percent in boys. The differences between the sexes begin to level off after the first year. While the total brain weight is greater in boys, the cerebellum of girls remains relatively larger. This relative size has been related to greater coordination, grace and delicacy in feminine motility.
An extensive series of studies which, due to their clarity and implications will probably become classic, are appropriately reported here. Freedman and Freedman (1974) observed Chinese-American (C-A) and European-American (E-A) infants while in a Hawaiian hospital nursery. The babies were observed for many categories of behavior: temperament, sensory development, autonomic and central nervous system maturity, motor development, social interest and response. A multivariate analysis of variance indicated that, while there was considerable overlap in range on all scales between the C-A and E-A infants, the two groups were decidedly and significantly different (p=.008). To cite two examples: (1) the typical supine E-A infant immediately struggled to remove a cloth placed firmly over his face; to the same stimulus, the C-A infant typically lay impassively; and (2) when placed in a prone position, the C-A infant lay as placed with face flat against the bedding whereas the E-A infant vigorously turned his face or lifted his head (there was no difference in ability to move the head between the groups). These measures indicate inherent (genetic) differences between the groups.
These investigators summarized their work by concluding that the two groups generally differed at birth in items which might be considered in the category of temperament. “The Chinese-American [infants] tended to be less changeable, less perturbable, tended to habituate more readily, and tended to calm themselves or to be consoled more readily when upset.” Other studies show these differences seem to characterize the groups at all later ages as well.
One might consider that these differences are due to other than genetic causes, e.g., nutrition or maternal temperament during gestation (embryonic environment). This is an unlikely explanation, however. On the one hand, all babies were selected from middle or higher socioeconomic class mothers with good diets. On the other hand, Navajo neonates in Arizona, studied by these same investigators, have essentially the same basic genetic stock as the Cantonese Chinese and show the same behavioral contrast with E-A infants. These behavioral dispositions at birth seem to persist throughout life and set the biases with which the individual will interact with his world.
Sexually dichotomous behaviors in all cultures become increasingly apparent throughout childhood so that between birth and puberty qualitative as well as quantitative pattern differences begin to manifest themselves. These reflect a basic sexual organization which is constitutionally biased. For example, the typical sex-related behaviors mentioned above hold for normal children, but girls subjected to androgenic influences during gestation, either from a malfunctioning adrenal or due to iatrogenic involvement, in comparison with normally developing girls and despite typical female upbringing, routinely show “tomboy” behavior in gross motor and muscular activity, and demonstrate choice preference for boys’ toys and games, together with a lack of interest in typically feminine pursuits such as infant care, playing house and being a homemaker in adulthood. On the other hand, males or females who are deprived of androgenic stimulation or exposed to excessive estrogens during gestation or whose tissues cannot respond to androgens, develop with feminoid behavioral traits regardless of how they are reared.
An extreme example in the other direction is seen with XY individuals who, due to a genetic trait, cannot respond to testosterone, crucial for normal male differentiation and development. These so-called “testicular females” or “androgen-insensitive” males display behavior traits which might be considered “hyper-female” despite typical female upbringing. These females almost invariably want children, a home life rather than careers, and when adopting children lavish them with attention and “mothering.” Without the ability to react to androgens they seem to have significantly few so-called male traits. Hermaphroditic and pseudo-hermaphroditic individuals whose rearing may be either male or female due to ambiguous genitalia and who adjust to the sex of assignment do so because associated with the condition is an accompanying biological flexibility unavailable to normal individuals (Diamond, 1968).
These social findings are predictable due to the biological inheritance of these individuals. In the normal population, of course, females with a typical upbringing may develop as “tomboys” and males may develop in a very passive effeminate mode; however, these occurrences are not abnormal and would not necessarily be predictable. The upbringing would be strongly effective in molding behavior in those children individuated to the respective patterns. That biology is crucially involved is apparent from the findings that, despite the aberrant hormone intervention, an androgenized female never is as fully masculinized as a male can be. Both genes and endocrines are mutually and interactionally involved with the environment, but the environment must work within the confines of biology.
While constitutional diasthetic biases exist, an individual’s behavior is modifiable by experience and environment. In a crude analogy: biology sets the sails and rudder, experience determines the direction and strength of the wind and tides. In normal development, positive reinforcement (reward) is evoked in the culture by the display of behaviors which are sex- (genital) appropriate, whereas those that are inappropriate either are not rewarded or are negatively reinforced. In all individuals, this conditioning process helps modify and shape behavior and, in complicated patterns such as the establishment and maintenance of social dominance or coitus, may even be necessary. There is no evidence, however, that such conditioning can completely structure behaviors such as are here discussed. In fact, by definition, learning is a modification of behavior rather than an initiation of original patterns. On the other hand, there are many clinical cases in which individuals are reared in a sex counter to their composite biology and yet, despite the social situation, the child strongly manifests his or her biological sex (Diamond, 1965). The conditioning processes hypothetically can be considered as evoking the neural circuitry related to the basic male and female patterns. The reward value of different reinforcements may, in its own way, be diasthetically organized by prior induction. For example, conditioning can strongly influence preferred games and activities, erotic situations, and partner types, but the greater male tendency and response to be aggressive and exploratory and the female tendency and preference to play in small groups, be nonaggressive and stay close to home is evolutionarily consistent. The environmental influences and upbringing must contend with these facts but can modify these patterns.
This does not mean that males and females cannot biologically be disposed other than as typically presented. It does mean, however, that when gender atypical behavior is seen, it might represent either an atypical biological development or a significantly strong experiential influence. Within certain constitutional diasthetic limits, individuals can be conditioned in any direction, but there may be a price to be paid if these limits are approached in a heterotypic direction. The price may be in a personal individual doubt of self-correctness (in performance, identity and choice of sexual partners). There is a growing body of data that show differential growth and development between boys and girls, and it is probable that the rate and extent of the neuro-physiological—including learning potential—, as well as general somatic developmental processes, underlying sex identification and object choice also differ between the sexes and thus mold sex-related behaviors.
Consider these signs of innate neural differences associated with learning and pattern acquisition that are manifest between birth and puberty. All such neural differences could influence perception and response capabilities. Neonatally, girls and boys visually fixate differentially to different targets. At 10 to 14 weeks of age, girls learn visual responses significantly better than boys when reinforcement is auditory and not visual. On the other hand, boys learn better when given visual reinforcement. Neonatal boys and girls differ in the amount of time they will visually fixate on a target object. Innate differences in response to, and use of, various sense modalities persist, as indicated by the well-known superiority of boys and adult males in visual-spatial skills. Males are significantly better than girls in adapting to glare and light stress. Girls, on the other hand, seem to have the advantage over boys in verbal and linguistic skills.
Many studies correlate well here. Two (Kimura, 1967; Taylor, 1969) have shown that the cerebral hemispheres of boys and girls develop differently with linguistic and auditory consequences. The right hemisphere develops earlier in females and this is supposedly the hemisphere more responsible for processing language; the left hemisphere is more crucial for speech detection. Lansdell (1962), by testing adults before and after brain surgery, has shown that males and females process visual and verbal material differently and in different hemispheres. He concludes that the physiological mechanisms underlying verbal ability and artistic judgment seem to overlap in the female but reside in opposite hemispheres in the male. Related to this are psychometric findings that girls show positive correlations between their level of artistic interest and their competence on verbal tests, while boys do not. Lansdell also, as others, noted sex differences related to short-term stress after which women are more relaxed than men, and men more vigilant.
The hemispheric myelination process itself appears to differ in boys and girls. This could no doubt account for some of the functional differences seen between the sexes. In the brains of 8 4-year-old children studied, myelination was greater in girls in the left-hand area of the motor cortex than in the motor cortex on the right. The reverse was true for boys.
Boys and girls show striking developmental differences in tactile thresholds in relation to dominant and nondominant hemispheres. Girls show significantly greater sensitivity on the dominant hand at age 5, with sensitivity changing to the nondominant hand, the adult pattern, at age 6. The boys show no significant difference in tactile sensitivity between their hands until age 11 when the nondominant hand becomes tactilely more sensitive.
Other neural differences between the sexes are manifest during infancy and childhood. Neonatal girls are more sensitive to general tactile stimuli, blanket removal, and airdraft stimulation than boys (they cry more quickly to these stimuli). The sexes, even cross-culturally, regularly differ in the age at onset of smile response.
In studies of 6-month-old children, girls, significantly more often than boys, showed visual interest fixation to pictures of human faces. Girls begin to smile before boys, and they are consistently easier to provoke to smile. As they develop they show greater interest in personal contact and a greater need for approval than do boys. These traits are believed to influence as well as be molded by culture. (Girls, by their smiles, may attract more attention or vice versa.)
As adults, more men than women report erotic arousal (reinforcement?) by visual stimuli. The auditory acuity of women is significantly greater than that of men, and women, more than men, seem to be aroused by “sweet talk” and reading (talking to the eye). To paraphrase an old saying, this may indicate that “the best way to a woman’s heart is through her ears.” (Interestingly, babies of both sexes respond preferentially to female voices—the higher pitches—than to male voices.)
Manifestation of cognitive and conative differences between boys and girls are seen in sexually different perceptual responses to Rorschach forms throughout the first 10 years of life, starting with codable verbal responses at 3 years of age. Boys give more responses in every category, see more movement, and make more mention of urogenital structures. Their comments tend to be more complicated, detailed, involved and rambling than those of girls, who respond more concretely and concisely with global responses.
As early as 7 months of age, boys show more anger, aggression and hostility than do girls. The development of fear reactions appears to be sex-linked. Males showing an early onset of fear during the first year of life show a significantly heightened fear reaction during their second year. This relationship does not exist for girls. Girls show more affection and friendliness than do boys. These differences persist to adulthood and are generally seen cross-culturally. These different social patterns also are seen consistently among many nonhuman species, thus following an evolutionary trend.
Sexual identity and object choice. From birth, with the assignment of a name and myriad other influences, individuals are subjected to environmental-social forces conducive to a specific sexual identity and choice of objects with which to interact. Since these will generally reinforce inherent tendencies, no difficulty will normally ensue. Typically, a boy will view himself as a boy, preferentially model after a male, and choose other boys as companions; vice versa for a girl. Society will encourage this. In some individuals, however, conflict (even where it is not yet recognized as such by the individual) may start prepubertally. In such children, discomfort in the assigned sex or toward certain objects or persons will be seen. For example, a child mistakenly brought up in a malassigned sex, or the one he or she prefers less, will often begin to expose this feeling of doubt or discomfort. The biological (neuroendocrine) forces attendant to identity and object choice for such individuals may not be fully active prepubertally, however, so the strength of conviction and ambivalence will not necessarily lead to socially unacceptable levels of demonstration.
Sexual identity, as the term is used here, refers to the individual’s personal and private assessment of his or her gender. Object choice, too, has a private and personal dimension. Sexual gender identity and object choice, publicly expressed in word or behavior by an individual, is comparable to his gender role. The gender role is stipulated by society, the identity is what the individual accepts for him or herself. In retrospect, despite the public gender role, many homosexuals and transsexuals (individuals in which sexual identity and object choices are called into question) report feelings of conflict occurring as early as 6 to 10 years of age. Among these groups will be many who, during these early years, displayed patterns more typical of members of the opposite sex than of their own. This public display is significant but not essential as a prognosticator of personal ambivalence.
Particularly revealing here are studies of female children who, during their prenatal and early postnatal life, were subject to abnormal amounts of androgenic hormones (those with the adrenogenital syndrome). These girls, significantly more often than normal girls, prefer to play with boys’ toys, have an aversion to, or are indifferent to, the care of babies, engage more often in intense outdoor activities, and are ambivalent toward being a girl or expressly desire to be a boy. These girls express less interest in future marriage and more interest in a career than do normal girls (Ehrhardt, Epstein, and Money, 1968).
To be sure, in the period between birth and puberty, the environment strongly affects the sexual development of the individual. Environmental influences serve, and in fact are needed, to elicit those behavioral characters programmed and allowable biologically. For example, a potential swimmer, musician, or copulator must have the opportunity to develop his or her ability and learn ways to maximize her or his potential. The extent to which the development or learning will occur will depend on the innate capacity of the individual to perform the particular behaviors at issue, as well as the ability of the individual to learn and practice. The flexibility of the dominant and subdominant sets of neural structures, and their sexual orientation, will determine how nurture and nature interact to influence human sexuality. It is likely that individuals with various types of biological anomalies of sexual development, e.g., pseudohermaphrodites, gain from the anomaly a flexibility and range of sexual dispositions much greater than found in normal persons. This flexibility would allow for a great deal of conditionability so that, regardless of conflicts between basic genetic sex and upbringing, the individual could accept his sex of rearing. This is possibly because prenatally the genetic-endocrine influences from one sex alone do not dominate. Simultaneously, this flexibility increases the extent to which an individual, in our gender oriented society, can gain from a strong, unisexual, unambiguous heritage.
Starting early in life, certainly much before the onset of puberty, a normal individual, to a limited degree, manifests levels of behavioral sexuality appropriate and specific for a male or female. Sexual patterns, a sense of sexual identity and bias toward certain sexual object choices, and genital mechanisms gradually develop and compose what passes for the individual’s personality and temperament. If these personality facets are manifest in a cultural manner appropriate to the morphological sex, all will go well. If, on the other hand, discrepancies exist the “environment” (e.g., parents or society) may become hostile, concerned or solicitous to the child and further complicate the developing process. In a hostile environment, the individual, too, may react in unpredictable ways.12 Despite a great deal of environmental input, the sexual personality of most individuals remains fairly stable from childhood to adulthood. Allowed to develop without undue restraint, no sudden changes in personality really occur. It might be good to repeat that generalities are being discussed. A great deal of individual variation exists.
Any complete discussion of sexual development should cover puberty and continue the theme through adulthood, maturity and into senescence. Sexual development is a continual process and predictable changes are seen to accompany all stages. For the present discussion, however, we will stop here, having shown how the prenatal processes are continued postnatally in a natural sequence, the early influences of induction affecting subsequent behavioral development.
As a last consideration, the following types of clinical findings are believed significant in exemplifying the power of inductive biological forces in organizing sexual identity and object choice in the face of conflicting environmental forces. Imperato-McGinley et al. (1974) report that in a small community in the Dominican Republic, due to a familial genetic-endocrine problem a large number of males were born which, at birth, appeared to be females. These males had bilateral testes as inguinal or labial masses, a labia-like scrotum, a urogenital sinus with a blind vaginal pouch, and a clitoris-like phallus. These males were raised as girls, yet:
Psychosexual orientation (post-pubertal) is male, and this isof considerable interest, since the sex of rearing in 18 of the affected males was female. Despite the sex of rearing, the affected were able to change gender identity at the time of puberty. They consider themselves as males and have a libido directed toward the opposite sex. Thus, male sex drive appears to be endocrine related, and the sex of rearing as female appears to have a lesser role in the presence of two masculinizing events, testosterone exposure in utero and again at puberty with the development of a male phenotype.
These individuals had obviously been so induced and organized biologically as males that their rearing, from birth to puberty as females, was not able to modify their gender identity nor object choice.
Transsexuals are individuals that as a group demonstrate a similar phenomenon. Although raised in one sex, these persons feel a “mistake in nature has been made” and request surgical alterations of their body to change to the opposite sex (see Benjamin, 1966). There is usually no evidence of an aberrant upbringing in the history of these individuals to account for this conflict between assigned sex and personal gender identity. In a similar vein, most homosexuals and heterosexuals seem to have been offered similar developmental environments and opportunities but develop differently in their object choice.
Lastly, in addition to those mentioned earlier, several groups of medically recognized individuals exist that are notable because their sexual behavior is incongruous with their upbringing but in keeping with their genetic-endocrine heritage. Such categories include: XXY (Klinefelter’s syndrome ) individuals who are phenotypic males, raised as males but usually show feminoid traits and atypically low libido; XO (Turner’s syndrome) individuals who are phenotypic females, raised as females and also showing atypically low libido; and boys born to mothers who received high doses of estrogen during pregnancy which induced extreme feminine behavior patterns despite normal upbringing as boys (Yalom, Green, and Fisk, 1973).
It is postulated that all types of heterosexual, homosexual and transsexual individuals are induced and biased by such biological forces. The environment in which an individual finds himself or herself will provide exposure to ways of modifying the sexual patterns or gender roles which he or she displays so as to feel most comfortable; it will do relatively less to the biases involved in gender identity and object choice. Each individual has an innate “feel” for what is right or comfortable sexually. The experiences of rearing are believed to provide enough different ways of achieving comfort within societally acceptable bounds. Nevertheless, enough data exist to indicate that even when society does not approve of certain sexual behaviors they will be manifest. The most rigid innate biases in this regard are believed to be in the area of gender identity and object choice. Clinical attempts in the use of psychoanalytic or aversion therapy to change extreme identity (e.g., transsexualism) and object choice (e.g., homosexuality) preferences have been routinely unsuccessful.13
Maccoby and Jacklin (1974) provide a great service by reviewing much of the literature relative to psychosexual and sociosexual development. They ask some highly significant questions and draw important conclusions:
Our analysis of the arguments concerning the role of modeling in sex-typing and our review of the research on selective imitation has led us to a conclusion that is very difficult to accept, namely that modeling plays a minor role in the development of sex-typed behavior. This conclusion seems to fly in the face of common sense and to conflict with many striking observations of sex-typed role playing on the part of children (p. 300).
In any case, existing evidence has not revealed any consistent process of “shaping” boys and girls toward a number of behaviors that are a normal part of our sex stereotypes (p. 348.)
It is reasonable then to talk about the process of sex-typed behavior—the learning of sex-typed behavior as a process built upon biological foundations that are sex-differentiated to some degree (p. 364.)
To interpret the findings of sexual development, they, as do others, consider the process of cognitive development as most explicitly formulated by Kohlberg (1966). This orientation postulates the child as organizing his or her life to conform to “cognitive organization of social-role concepts around universal physical dimensions which accounts for the existence of universals in sex-role attitudes. [The] theory then is cognitive in that it stresses the active nature of the child’s thought as he organizes his role perceptions and role learnings around his basic conceptions of his body and his world (Kohlberg, 1966, pp. 82-83).” To Kohlberg’s ideas, Maccoby and Jacklin add importance to direct reinforcement and imitation. I would simply say that it is likely that the genetic and endocrine forces to which this chapter points are involved in the organization of cognitive development processes, reinforcement qualities and predispositions to imitate.14
SUMMARY
In this chapter, I have tried to trace the early development of sexual behavior capacity during the life span of an individual. The development of biological and behavioral potentials is seen to be related and thus have ramifications strongly affecting theories which relate to childrearing and adult sexuality.
In this discussion I have introduced three concepts which I believe will prove fruitful in dealing with some of the most mystifying anomalies of development, such as the way in which individuals can apparently be inconsistent or discordant in their expression of sexuality across various parameters of consideration. The first concept emphasizes a parallel between the simultaneous development of different structures and physiological mechanisms used in sexual and reproductive activities, and different behavioral patterns which would be associated with these same processes.
Second, I consider that a single continuum does not provide an adequate model of masculine or feminine sex behavior development. In many ways both sexes develop or could develop similarity. To distinguish the sexes, however, one must consider separate criteria for male and female, and masculinity or femininity, not put them as poles on one continuum. Further, these dichotomous comparisons must be considered for reproductive functions and a minimum of four behavioral areas which comprise a basic sexual behavior profile for an individual: patterns, mechanisms, identity and object choice. The combinations and permutations of these parameters of sexuality are numerous enough to provide a broad spectrum of sexuality, especially when learning effects are added. Seemingly, this reflects an unfathomable admixture. Actually, these should be seen as indicating the extreme flexibility inherent in human sexual behavior.
Third, I have presented a concept of differing rates or susceptibilities of development among neural tissues mediating these behavioral areas and morphology, to provide a framework within which the etiology of transsexualism, homosexualism and heterosexualism can be equally considered.
Our culture now is involved with a strong concern with the societal stereotypes of gender roles and their inflexibility. The generalities of behavior and its biological basis presented in this chapter must be considered with the constant memory that the variability for each category is large, and each individual always must be judged in his or her own right. A goal for our culture might be to recognize and accept that generalities can exist simultaneously with allowable deviation from the typical. This allowance would permit each individual sufficient freedom to develop and express his or her potential in any direction, on all levels of sexuality, without attaching a negative value to any variation just because it is different. We can, however, continue to explore what factors produce these differences.